Влияние АКТГ на долговременную память
Влияние АКТГ на долговременную память также зависит от дозы и времени его введения. Так, по данным R. L. Cold и К. В. van Buskirk (1976), введенный в дозе 0,03—0,3 ME, он увеличивал время сохранения пассивного навыка избегания у крыс, а в дозе 3—10 ME вызывал ретроградную амнезию Стимулирующие эффекты АКТГ независимо от дозы проявлялись сильнее при обучении и проверке выработанного навыка в вечерние часы, а также при повышении мотивационного значения аверсивного стимула, а уменьшались по мере увеличения силы раздражающего электрического тока и интервала времени между окончанием обучения и инъекцией вещества.
По-видимому, стимулирующее влияние АКТГ на процессы хранения энграмм долговременной памяти связано с его избирательным влиянием на процессы консолидации.
Многие исследователи отводят значительную роль в механизме его действия эмоциональной значимости подкрепляющих факторов, поскольку влияние АКТГ на память проявляется наиболее отчетливо в экспериментах с отрицательным подкреплением. Более того, P. Gray (1975) и другие авторы полагают, что влияние АКТГ на скорость выработки навыка условного избегания у крыс связано с формированием особого состояния при обучении (state, dependent learning).
В частности, АКТГ улучшал воспроизведение этого навыка у крыс, получивших его при обучении, но не изменял его в тех случаях, когда при обучении животным вместо АКТГ вводили физиологический раствор.
Возможно, что способность АКТГ увеличивать время сохранения информации в ЦНС связана с кортикостероидами надпочечников. Действительно, кортикостерон, гидрокортизон и дексаметазон [Flood J. F. et al., 1978] также увеличивали время хранения закрепленных навыков активного и пассивного условного избегания у мышей. Они также замедляли процесс угашения пищедобывательных и оборонительных условных рефлексов [Митюшов М. И. и Соколова Е. В., 1974; Hennessy J. W. et al., 1973].
Более того, дексаметазон устранял амнезию, вызванную анизомицином, — ингибитором синтеза белка [Flood J. F. et al., 1978]. H. Righter (1975) рассматривает уровень кортикостерона в плазме при введении АКТГ4-10 как показатель уменьшения амнезии, вызванной углекислым газом у крыс.
Приведенные данные позволяют сделать вывод об общем регуляторном стимулирующем воздействии гипофизарноадреналовой системы (АКТГ и кортикостероидов) на процессы формирования и длительного хранения энграмм долговременной памяти у животных и человека. Однако это предположение оспаривается A. J. Dunn и Н. Liebman (1977) на том основании, что аминоглутетимйд, тормозящий секрецию кортикостероидов в ответ на введение АКТГ, не вызывал у мышей амнезии выработанного навыка пассивного избегания электрошока.
Кроме того, введение анизомицина и циклогексемида, вызывающих амнезию, снижало уровень кортикостерона у обученных животных лишь в первые 20 мин, а через 2 ч после инъекции секреция кортикостерона повышалась в 5 раз [Flood J. F. et al., 1978]. Таким образом, гипотеза об общем регуляторном стимулирующем воздействии гипофизарноадреналовой системы на различные процессы памяти требует дальнейшей эксперимен тальной проверки.
«Нейрохимические и функциональные основы долговременной памяти»,
Ю.С. Бродкин, Ю.В. Зайцев

Многие из этих пептидов находятся в нервных окончаниях и транспортируются к синапсам с помощью аксоплазматического тока. Субклеточное распределение нейротензина, соматостатина, ангиотензина, субстанции Р, вазопрессина и окситоцина характеризуется их преимущественным связыванием с синаптосомами, выделенными из различных внегипоталамических образований головного мозга [Larner J., 1977; Kobayashi R. M. et al., 1977; Bury R. W. et al., 1977]. Это…
Продукты ферментативного расщепления вазопрессина и окситоцина со специфической последовательностью аминокислот вызывали деполяризацию идентифицированных нейронов гигантской улитки [Takeuchi H. et al., 1977] Тиреотропин-рилизинг-фактор (тиреолиберин) и рилизингфактор лютеинизирующего гормона (люлиберин), а также соматостатин уменьшали спонтанную импульсную активность нейронов коры головного мозга и мозжечка, ствола мозга и гипоталамуса у крыс при ионофоретическом введении. Эти пептидные гормоны уменьшали возбуждение…
Учитывая, что пептидные гормоны существенно изменяют уровень возбудимости нейронов и нейронных популяций в различных образованиях головного мозга, представляло значительный интерес установить зависимость между этими эффектами и влиянием пептидных гормонов на обучение, фиксацию и хранение информации в мозге животных и человека. Больше всего экспериментов проведено по изучению влияния на эти процессы АКТГ. Еще в 1969 г….
Сходное с АКТГ влияние на память и обучение животных оказывает α-меланоцитстимулирующий гормон, структура которого близка к отдельным фрагментам АКТГ: он ускоряет обучение [Paterson А. Т., 1975], значительно увеличивает время хранения закрепленных навыков активного и пассивного избегания электрошока у крыс и облегчает их воспроизведение [Веckwitt В. Е. et al, 1977]. Нужно отметить, что АКТГ, его отдельные…
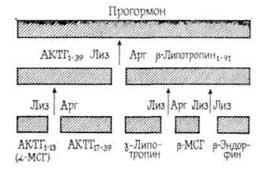
Необходимо отметить, что активным влиянием на память обладают не целые молекулы пептидных гормонов, а в основном их отдельные фрагменты, характеризующиеся специфической последовательностью аминокислот. Следовательно, можно предположить, что эти фрагменты образуются в организме в процессе активного метаболизма пептидных гормонов под влиянием протеолитических ферментов [Чипенс Г. И. и др., 1975]. Интимный механизм пептидных гормонов окончательно не выяснен,…