Продукты ферментативного расщепления вазопрессина и окситоцина
Продукты ферментативного расщепления вазопрессина и окситоцина со специфической последовательностью аминокислот вызывали деполяризацию идентифицированных нейронов гигантской улитки [Takeuchi H. et al., 1977] Тиреотропин-рилизинг-фактор (тиреолиберин) и рилизингфактор лютеинизирующего гормона (люлиберин), а также соматостатин уменьшали спонтанную импульсную активность нейронов коры головного мозга и мозжечка, ствола мозга и гипоталамуса у крыс при ионофоретическом введении. Эти пептидные гормоны уменьшали возбуждение нейронов, вызванное глутаминовой кислотой, и вызывали гиперполяризацию мембраны нейронов [Renaud L. Р. et al., 1978].
Большой интерес представляют исследования J. L. Barker (1978) по изучению влияния пептидов на нейрональную мембрану, выполненные на нейронах апплизии и улитки, а также культуре нейронов спинного мозга. Автор использовал субстанцию Р, лейэнкефалин, вазопрессин и окситоцин. Оказалось, что для последних были характерны четыре типа эффекта.
Во-первых, эффекты, сходные с действием возбуждающих или тормозящих нейромедиаторов. Отличие заключалось лишь в том, что гиперполяризация, вызываемая этими пептидами, не проявляла тенденции к десенситизации, тогда как деполяризация имела такую тенденцию на фоне повышения мембранного сопротивления. Второй эффект действия пептидов проявлялся внезапным угнетением возбудимости нейронов за счет повышения порога возникновения.
Эффект третьего типа обусловлен способностью усиливать или ослаблять влияние классических нейромедиаторов на мембрану нейронов (в частности, энкефалин подавляет ответ нейрона на глутамат и усиливает деполяризацию его мембраны, обусловленную глицином). Эффект четвертого типа определяется способностью пептидов резко снижать мембранный потенциал. При отмывании пептида или прекращении ионофореза его действие легко исчезает. К. Wishnewski (1979) считает, что влияние на мембрану нейрона является общим свойством всех пептидов.
Нами [Вислобоков А. И. и Зайцев Ю. В., 1979] изучено влияние одного из фрагментов АКТГ (АКЛТ4-10) на мембрану идентифицированных нейронов прудовика.
Оказалось, что этот фрагмент незначительно изменяет мембранный потенциал, вызывая небольшую деполяризацию мембраны, однако он в 2—3 раза увеличивает амплитуду ВПСП, несколько снижая их частоту. Возможно, что эффект АКТГ обусловлен его пресинаптическим облегчающим влиянием на высвобождение нейромедиатора, не связанным с непосредственным воздействием на мембрану нейрона.
«Нейрохимические и функциональные основы долговременной памяти»,
Ю.С. Бродкин, Ю.В. Зайцев

Многие из этих пептидов находятся в нервных окончаниях и транспортируются к синапсам с помощью аксоплазматического тока. Субклеточное распределение нейротензина, соматостатина, ангиотензина, субстанции Р, вазопрессина и окситоцина характеризуется их преимущественным связыванием с синаптосомами, выделенными из различных внегипоталамических образований головного мозга [Larner J., 1977; Kobayashi R. M. et al., 1977; Bury R. W. et al., 1977]. Это…
Учитывая, что пептидные гормоны существенно изменяют уровень возбудимости нейронов и нейронных популяций в различных образованиях головного мозга, представляло значительный интерес установить зависимость между этими эффектами и влиянием пептидных гормонов на обучение, фиксацию и хранение информации в мозге животных и человека. Больше всего экспериментов проведено по изучению влияния на эти процессы АКТГ. Еще в 1969 г….
Влияние АКТГ на долговременную память также зависит от дозы и времени его введения. Так, по данным R. L. Cold и К. В. van Buskirk (1976), введенный в дозе 0,03—0,3 ME, он увеличивал время сохранения пассивного навыка избегания у крыс, а в дозе 3—10 ME вызывал ретроградную амнезию Стимулирующие эффекты АКТГ независимо от дозы проявлялись сильнее…
Сходное с АКТГ влияние на память и обучение животных оказывает α-меланоцитстимулирующий гормон, структура которого близка к отдельным фрагментам АКТГ: он ускоряет обучение [Paterson А. Т., 1975], значительно увеличивает время хранения закрепленных навыков активного и пассивного избегания электрошока у крыс и облегчает их воспроизведение [Веckwitt В. Е. et al, 1977]. Нужно отметить, что АКТГ, его отдельные…
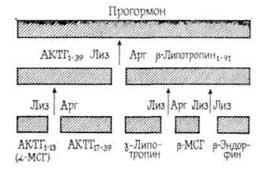
Необходимо отметить, что активным влиянием на память обладают не целые молекулы пептидных гормонов, а в основном их отдельные фрагменты, характеризующиеся специфической последовательностью аминокислот. Следовательно, можно предположить, что эти фрагменты образуются в организме в процессе активного метаболизма пептидных гормонов под влиянием протеолитических ферментов [Чипенс Г. И. и др., 1975]. Интимный механизм пептидных гормонов окончательно не выяснен,…