Изменения уровня возбудимости нейронов и нейронных популяций в различных образованиях головного мозга
Учитывая, что пептидные гормоны существенно изменяют уровень возбудимости нейронов и нейронных популяций в различных образованиях головного мозга, представляло значительный интерес установить зависимость между этими эффектами и влиянием пептидных гормонов на обучение, фиксацию и хранение информации в мозге животных и человека. Больше всего экспериментов проведено по изучению влияния на эти процессы АКТГ.
Еще в 1969 г. D. de Wied показал, что после удаления гипофиза животные не только утрачивают способность к обучению, но и ранее выработанные навыки. Так, дефицит навыка активного избегания у крыс можно было устранить не только введением АКТГ, вазопрессина или амелано-цитстимулирующего гормона, но и отдельными фрагментами этих пептидов, утратившими способность регулировать секрецию кортикостероидов и другие гормонрегулирующие функции. Впоследствии D. de Wied и сотр. (1975, 1977, 1979) показали, что аналоги АКТГ способствуют не только сохранению навыка активного избегания, но и устойчивости зрительных дифференцировок.
Они замедляют угашение инструментальных пищедобывательных реакций и навыка пассивного избегания электрошока, облегчают переобучение, устраняют ретроградную амнезию, вызванную углекислым газом.
J. Verhoff и соавт. (1977) изучали зависимость между структурой аналогов АКТГ и их способностью закреплять в памяти навык условного избегания электроболевого раздражения у крыс. Оказалось, что наибольшей активностью обладает фрагмент АКТГ4-Э— гексапептид, у которого а-изомер аргинина в положении 8 заменен D-изомером лизина.
Наиболее подробно исследованы лево и правовращающие изомеры АКТГ в пределах участка 4—10 его аминокислотной последовательности.
Влияние различных факторов АКТГ на длительность сохранения навыка активного избегания электроболевого раздражения у крыс (de Wied D. et al., 1975)
| Фрагмент АКТГ | Последовательность аминокислот | Степень активности |
| АКТГ4-10 | Н-Мет-Глу-Гис-Фен-Арг-Трп-Гли-ОН | + |
| АКТГ4-7 | Н-Мет-Глу-Гис-Фен-ОН | + |
| 7Д-Фен-АКТГ4-10 | Н-Мет-Глу-Гис-дФен-Арг-Трп-Гли-ОН | — |
| 4Д-Мет-8-Лизин-АКЛТ4-10 | Н-дМет-Глу-Гис-Фен-Лиз-Трп-ОН | ++ |
| АКТГС6-9 | Н-Гис-Фен-Арг-Трп-ОН | 0 |
| 5Д-Глу-8-Лизин-АКТГ4-9 | Н-Мет-дГлу-Фен-Лиз-Трп-ОН | ++ |
| 8Д-Лизин-АКТГ4-9 | Н-Мет-Глу-Гис-Фен-дЛиз-Трп-ОН | +++ |
| 9Д-Трп-8-Лизин-АКТГ4-9 | Н-Мет-Глу-Гис-Фен-Лиз-дТрп-ОН | ++ |
| 4-Мет(0)-8-Лизин-АКТГ4-9 | Н-Мет-Глу-Гис-Фен-Лиз-дТрп-ОН | + |
| 4-Мет(0)-8Д-Лизин-9-Фенил-АКТГ4-9 | Н-Мет(0)-Глу-Гис-Фен-дЛиз-Фен-ОН | +++++ |
Примечание: (+) — торможение угашения навыка; (—) — облегчение угашения; (0) — отсутствие эффекта.
Значительной активностью обладают и другие фрагменты, в частности АКТГ4-10 и АКТГ4-7. Однако у АКТГ6-9 она резко ослаблена и, по данным R. L. Isaacson с соавт. (1976), практически отсутствует у АКТГ57 (Н-глугис-фен-ОН).
Как известно, АКТГ при системном введении не проникает через гематоэнцефалический барьер, и поэтому его влияние на высшие функции ЦНС связывают с образованием вышеперечисленных фрагментов, которые захватываются из тока крови тканью мозга и ретроградно транспортируются через гипофиз и базальный гипоталамус. Согласно многочисленным данным [Bonus В., 1979, и др.], наиболее выраженным влиянием на память обладает фрагмент АКТГ4-10, который имеется также в молекулах и β-меланоцитстимулпрующих гормонов и β-липотропина (фрагмент 47—53).
Для него характерна кратковременность действия на поведение (до нескольких часов).
По данным F. С. Colpaert с соавт. (1978), АКТГ4-10 усиливает внимание, активирует механизмы по оценке информации на уровне сенсорного входа. Влияние этих пептидов на высшие функции проявляется в усилении мотиваций, внимания, облегчении обучения.
«Нейрохимические и функциональные основы долговременной памяти»,
Ю.С. Бродкин, Ю.В. Зайцев

Многие из этих пептидов находятся в нервных окончаниях и транспортируются к синапсам с помощью аксоплазматического тока. Субклеточное распределение нейротензина, соматостатина, ангиотензина, субстанции Р, вазопрессина и окситоцина характеризуется их преимущественным связыванием с синаптосомами, выделенными из различных внегипоталамических образований головного мозга [Larner J., 1977; Kobayashi R. M. et al., 1977; Bury R. W. et al., 1977]. Это…
Продукты ферментативного расщепления вазопрессина и окситоцина со специфической последовательностью аминокислот вызывали деполяризацию идентифицированных нейронов гигантской улитки [Takeuchi H. et al., 1977] Тиреотропин-рилизинг-фактор (тиреолиберин) и рилизингфактор лютеинизирующего гормона (люлиберин), а также соматостатин уменьшали спонтанную импульсную активность нейронов коры головного мозга и мозжечка, ствола мозга и гипоталамуса у крыс при ионофоретическом введении. Эти пептидные гормоны уменьшали возбуждение…
Влияние АКТГ на долговременную память также зависит от дозы и времени его введения. Так, по данным R. L. Cold и К. В. van Buskirk (1976), введенный в дозе 0,03—0,3 ME, он увеличивал время сохранения пассивного навыка избегания у крыс, а в дозе 3—10 ME вызывал ретроградную амнезию Стимулирующие эффекты АКТГ независимо от дозы проявлялись сильнее…
Сходное с АКТГ влияние на память и обучение животных оказывает α-меланоцитстимулирующий гормон, структура которого близка к отдельным фрагментам АКТГ: он ускоряет обучение [Paterson А. Т., 1975], значительно увеличивает время хранения закрепленных навыков активного и пассивного избегания электрошока у крыс и облегчает их воспроизведение [Веckwitt В. Е. et al, 1977]. Нужно отметить, что АКТГ, его отдельные…
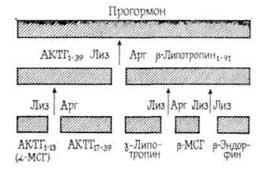
Необходимо отметить, что активным влиянием на память обладают не целые молекулы пептидных гормонов, а в основном их отдельные фрагменты, характеризующиеся специфической последовательностью аминокислот. Следовательно, можно предположить, что эти фрагменты образуются в организме в процессе активного метаболизма пептидных гормонов под влиянием протеолитических ферментов [Чипенс Г. И. и др., 1975]. Интимный механизм пептидных гормонов окончательно не выяснен,…