ЛДГ в цитоплазме культуры ФЭК, инфицированной онковирусом А12
В цитоплазме культуры ФЭК, инфицированной онковирусом А12, ЛДГ5 не найдена, а ЛДГ4 исчезала на 18-й и 24-й дни инфекции. В остальные дни активность фермента была меньше как по сравнению с нормой, так и по сравнению с активностью в клетках ФЭК, зараженной вирусом А6.
Активность ЛДГ1 существенно снизилась в цитоплазме на 24-й день (в 1,6 раза), а ЛДГЗ уменьшилась в 1 1/2 раза на 18-й день и 1,3 раза на 24-й день как по сравнению с нормой, так и с активностью в клетках ФЭК, зараженной А6. В ядерной фракции на 5-й и 18-й дни после инокуляции вируса А12 в культуру ФЭК было обнаружено достоверное увеличение активности МДГ1, активность МДГ2 повышалась на 3-й, 5-й и 8-й дни, а МДГЗ — на 3-й и 24-й дни.
В цитоплазме под действием онковируса активность МДГЗ возрастала на 3-й и 24-й дни инфекции.
Таким образом, действие инфекционных и онкогенных вирусов вызывает перестройку в изоферментных спектрах ферментов энергетического обмена как в ядерной фракции, так и в цитоплазме.
Эти нарушения обнаруживаются с первых дней взаимодействия клеток культуры ФЭК с вирусом. Ори контакте с онкогенным вирусом ОЙИ выражены больше, чем при контакте с инфекционным.
Повышение активности некоторых ферментов энергетического обмена в цитоплазме (по сравнению с ядерной фракцией) при заражении инфекционным и онкогенным вирусами, вероятно, можно объяснить более высоким уровнем гликолитических процессов, стимулированных онковирусом в трансформированных клетках. Важно подчеркнуть, что эти сдвиги в регуляции синтеза изоформ исследованных ферментов не связаны с пролиферативным ростом культуры клеток, а обусловлены именно воздействием вируса на клетку. Возможно, они происходят вследствие эпигеномной перестройки под влиянием онковируса.
Особый интерес представляют данные сравнительного анализа сдвигов в изоформах исследуемых ферментов в культурах клеток ФЭК, инфицированных вирусом А12, и в клетках сарком А12 и ретикулосарком 321-КРС, растущих in vitro.
Установлено, что в клетках саркомы А12 в ядерной фракции преобладает гексокиназа 3. Общая активность фермента в цитоплазме клеток была выше, чем в ядре. Аналогичные сдвиги отмечены нами и в процессе трансформации клеток ФЭК, инфицированных этим же вирусом.
Смотрите рисунок — Активность ПК
Смотрите рисунок — Электрофореграммы и денситограммы изоферментов ГК (1 — 3) в глобулиновой фракции ядер клеток культуры ФЭК
Изоферменты ГК (1 — 3) в саркоме хомяка, индуцированной аденовирусом 12-го типа
 в саркоме хомяка, индуцированной аденовирусом 12-го типа")
а — в ядерной фракции iv vitro; б — в системе in vivo; в — в цитоплазме in vivo.
Изоферменты ГК (1 — 3) в крысиной ретикулосаркоме КРС — 321
 в крысиной ретикулосаркоме КРС — 321")
а, б, в — то же, что на рисунке выше.
Электрофореграммы и денситограммы изоферментов ГК (1 — 3)
в саркоме хомяка, индуцированной вирусом А12
 в саркоме хомяка, индуцированной вирусом А12")
а — в растворимой ядерной фракции; 6 — в цитоплазме.
Изоферментный спектр ЛДГ в ядрах клеток саркомы А12 представлен двумя формами, из которых преобладала ЛДГ1. В цитоплазме выявлены 4 изофермента.
Изоферменты ЛДГ (1 — 4) в саркоме хомяка
 в саркоме хомяка")
В ядерной фракции системы; а — in vivo; б — in vitro; в цитоплазме; в — in vivo; г — in vitro.
Малатдегидрогеназа в ядре в цитоплазме клеток саркомы А12 представлена тремя изоферментами. В ядерной фракции их количество примерно одинаковое, а в цитоплазме преобладала МДГЗ.
Изоферменты МДГ (1 — 3) в саркоме хомяка
 в саркоме хомяка")
а — в ядерной фракции in vitro; б — в системе in vivo; в — в цитоплазме in vivo; r — в культуре.
Для ретикулосаркомы крыс 321-КРС общая активность гексокиназы в цитоплазме клеток была выше, чем в ядре (8,5 и 2,8 мкмоль).
В ядерной фракции преобладала гексокиназа 3.
Смотрите рисунок — Изоферменты ГК (1 — 3) в крысиной ретикулосаркоме КРС — 321
Электрофореграммы и денситограммы изоферментов ГК (1 — 3) в крысиной ретикулосаркоме КРС-321 in vivo
 в крысиной ретикулосаркоме КРС-321 in vivo")
а — в ядерной фракции; б — в цитоплазме.
Изоферментный спектр ЛДГ в клетках этой саркомы был представлен двумя изоферментами как в цитоплазме, так и в ядерной фракции. Активность каждого из изоферментов (ЛДП и ЛДГ2) в ядре была выше их активности в цитоплазме в культуре.
Изоферменты ЛДГ (1 — 2) в крысиной ретикулосаркоме КРС-321
 в крысиной ретикулосаркоме КРС-321")
В ядерной фракции системы: а — in vivo; б — in vitro; в цитоплазме: в — in vivo; г — in vitro.
Электрофореграммы и денситограммы изоферментов ЛДГ (1 — 2)
в крысиной ретикулосаркоме КРС-321 in vivo
 в крысиной ретикулосаркоме КРС-321 in vivo")
а — в ядерной фракции; б — в цитоплазме.
Малатдегидрогеназа в клетках саркомы 321-КРС выявлена в трех формах и в ядерной фракции, и в цитоплазме. В ядерной фракции содержание МДГЗ было значительно ниже, чем количество двух других изоферментов. В цитоплазме клеток преобладала МДГ2. Оказалось, что в ядерной фракции уровень МДГ1 и МДГЗ был выше, чем уровень этих изоферментов в цитоплазме клеток культур.
Изоферменты МДГ (1 — 3) в крысиной ретикулосаркоме КРС-321
 в крысиной ретикулосаркоме КРС-321")
а, б, в, г — то же, что на рисунке выше.
Смотрите рисунок — Изоферменты ЛДГ (1 — 2) в крысиной ретикулосаркоме КРС-321
Электрофореграммы и денситограммы изоферментов МДГ (1 — 3)
в крысиной ретикулосаркоме КРС-321 in vivo
 в крысиной ретикулосаркоме КРС-321 in vivo")
а, б — то же, что на рисунке выше.
Смотрите рисунок — Электрофореграммы и денситограммы изоферментов ЛДГ (1 — 2) в крысиной ретикулосаркоме КРС-321 in vivo
Таким образом, исследование в динамике молекулярных форм ферментов энергетического обмена в культуре, зараженной онкогенным аденовирусом 12-го тина, и в саркомах А12 и 321-КРС позволило выявить общие черты в изменении каждого из изученных ферментов. Наиболее контрастными были изменения гексокиназы. Прежде всего они выражались в повышенном содержании фермента в цитоплазме по сравнению с его уровнем в ядре клеток культуры ФЭК при появлении очагов морфологической трансформации.
Отмечено преобладание гексокиназы 3 над другими изоферментами в ядерной фракции. Идентичные результаты получены и на других тест-системах. Так, значительное повышение относительного содержания гексокиназы 3 в клетках гепатомы Морриса коррелировало с увеличением степени злокачественности исследованных новообразований. Этот изофермент был преобладающим также в карциномах молочной железы, почки, в рабдомиосаркоме человека (В. С. Шапот, 1975).
Гексокиназе придается особое значение, так как предполагают, что в поверхностных мембранах раковых клеток локализована гексокиназа с повышенным сродством к глюкозе, вероятно, преимущественно этот изофермент выходит в окружающую среду.
Возможно, увеличение содержания гексокиназы, обеспечивающее повышение потенциальной способности к гликолизу, связано с малигнизацией клеток. В ядрах клеток ФЭК, зараженных онкогенным вирусом А12, спектр изоформ лактатдегидрогеназы более упрощен, чем в цитоплазме, за счет быстро мигрирующих изоферментов. Малатдегидрогеназа в этих же клетках представлена тремя формами как в ядре, так и в цитоплазме. В обеих фракциях содержание МДГЗ было повышено. Аналогичные изменения спектров ЛДГ и МДГ наблюдались и в культуре клеток сарком А12.
Таким образом, изменения изоферментных спектров гексокиназы, ЛДГ и МДГ в растворимой ядерной фракции и цитоплазме клеток ФЭК под влиянием ДНК-содержащего онковируса свидетельствуют об усилении гликолитических процессов, которые характерны для малигнизированной ткани.
Следовательно, обнаруженные сдвиги можно использовать в качестве объективных признаков трансформации клеток. Предполагают, что повышение активности гексокиназы и стимуляция гликолиза на ранних стадиях трансформации клеток ФЭК под действием онковируса являются основным свойством бластомогенного процесса, возможно, обусловленным нерегулируемостью процесса пролиферации клеток.
«Механизмы вирусного онкогенеза»,
А.И.Агеенко
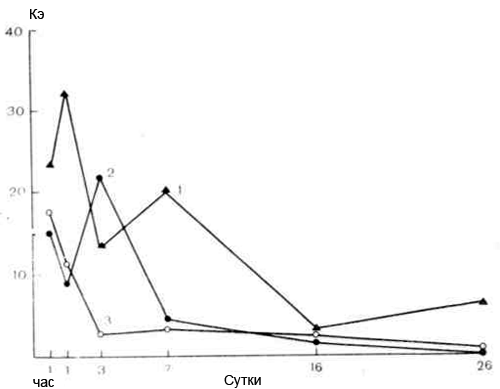
Анализируя результаты, полученные при количественном исследовании тонкого строения клеток ФЭХ при действии вируса А12, можно отметить, что показатель Кэ, характеризующий степень развития эргастоплазмы, значительно снижается, особенно начиная с 7-го дня после инфицирования, и к сроку выраженной трансформации достигает предельно низких величин. Динамика изменения показателя Кэ в процессе трансформации культуры ФЭХ вирусами А12 и RSV 1…
Особое значение для оценки сдвигов, происходящих в энергетическом обмене на ранних этапах трансформации, детерминированной онковирусом, приобретает анализ структурной перестройки митохондрий, осуществляемый параллельно с определением функциональных изменений, полученных на этой же системе и в те же сроки с помощью биохимических методов. Сведения о подобных исследованиях в литературе отсутствуют. А. И. Агеенко, А. С. Ягубов и Ю….
К принципиально важным вопросам онкоморфологии относятся выявление и определение сходства и отличий нормальных и опухолевых клеток. Это стало возможным только в последние годы благодаря количественной электронной микроскопии, основанной на морфометрическом анализе электронно-микроскопических картин с использованием методов планиметрии и принципов стереологии (Г. Г. Автандилов, 1973). Установлено, что эргастоплазма и полирибосомный аппарат являются основными цитоплазматическими органеллами белкового…
Преобладание медленно движущихся (катодных) фракций ЛДГ4 и ЛДГ5 при снижении уровня быстро движущихся (анодных) ЛДГ1 и ЛДГ2 обнаружено в тканях, адаптированных к гипоксии и ацидозу, в которых основным источником энергии является гликолиз. В то же время можно считать доказанным, что удельный вес гликолиза в клетках злокачественных новообразований повышается (В. С. Шапот, 1975). Установлено, что скорость…
Показано, что повышение процента ЛДГ4 и ЛДГ5, а также снижение содержания ЛДГ1 и ЛДГ2 наблюдались в тканях у большей части обследованных больных (Р<0,001), не только в гомогенатах злокачественных опухолей, но и в тканях полипов прямой и ободочной кишки. При этом отсутствие увеличения содержания ЛДГ5 в опухоли при раке выявлено только в 8,7% случаев (в 17…