Количественный анализ структурной перестройки эргастоплазмы и полирибосомного аппарата клеток в динамике вирусного онкогенеза
К принципиально важным вопросам онкоморфологии относятся выявление и определение сходства и отличий нормальных и опухолевых клеток. Это стало возможным только в последние годы благодаря количественной электронной микроскопии, основанной на морфометрическом анализе электронно-микроскопических картин с использованием методов планиметрии и принципов стереологии (Г. Г. Автандилов, 1973).
Установлено, что эргастоплазма и полирибосомный аппарат являются основными цитоплазматическими органеллами белкового синтеза.
Эргастоплазма имеет непосредственное отношение к синтезу высокоспециализированных белковых субстратов, а полирибосомный аппарат ответствен за синтез неспецифических белков, необходимых для роста и размножения клеточной массы. В процессе малигнизации происходит значительная перестройка большинства надмолекулярных систем клетки и в первую очередь белоксинтезирующего аппарата.
Эти данные получены при сравнительном анализе нормальных и опухолевых клеток. Работы же, посвященные изучению структурной перестройки клеток в динамике вирусного канцерогенеза, в доступной литературе отсутствуют.
В связи с этим значительный интерес представляло изучить специфику изменений строения эргастоплазмы и полирибосомного аппарата клетки в динамике вирусного онкогенеза, а также сравнить действия РНК- и ДНК-геномных вирусов на характер изменений указанных органелл, поскольку эти онкогенные агенты имеют различный генетический аппарат и различно взаимодействуют с клеткой.
Естественно, для решения поставленных задач целесообразно применение методов морфометрии, которые значительно объективизируют информацию о тонком строении органелл клетки.
А. С. Ягубов и А. И. Агеенко (1975) использовали первичную культуру фибробластов эмбриона хомяка (ФЭХ).
Эту культуру разделили на три серии. После формирования монослоя клетки инфицировали в одной серии вирусом саркомы Рауса штамм Шмидта—Руппина (1-я серия) и вирусом А12 (2-я серия), 3—6-я серии служили контролем. Исследование производили через 1—2 ч после заражения и далее через 1, 3, 7 сут, на 16-е сутки (начало видимой трансформации) и на 26-е сутки (выраженная морфологическая трансформация).
Для морфометрического анализа отбирали по 30 клеток из каждой серии.
По фотоотпечаткам (методику, смотрите ссылку ниже) определяли следующие морфометрические характеристики: площадь поверхности мембран эргастоплазмы (Sэv), объем эргастоплазмы (Vэv), число рибосом, прикрепленных на единице длины мембран эргастоплазмы (NрэL), коэффициент фрагментации эргастоплазмы (Кэф).
Вычисляли коэффициент уровня морфологической организации эргастоплазмы: Кэ =Sэv*Vэv*NрэL*1/Кэф.
Коэффициент развития полирибосомального аппарата определяли как Kп = Nрпа, т. е. по количеству рибосом, объединенных в полисомы на 1 мк2 цитоплазмы.
Кроме этого, исследовали отношение Кэ/Кп, отражающее соотношение экстра- и интрацеллюлярного белкового синтеза. Все морфометрические параметры и коэффициенты подвергались статистической обработке. Доверительные интервалы средних взяты с надежностью 95%.
«Механизмы вирусного онкогенеза»,
А.И.Агеенко
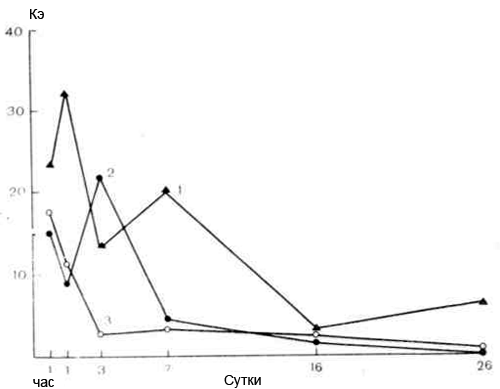
Анализируя результаты, полученные при количественном исследовании тонкого строения клеток ФЭХ при действии вируса А12, можно отметить, что показатель Кэ, характеризующий степень развития эргастоплазмы, значительно снижается, особенно начиная с 7-го дня после инфицирования, и к сроку выраженной трансформации достигает предельно низких величин. Динамика изменения показателя Кэ в процессе трансформации культуры ФЭХ вирусами А12 и RSV 1…
Особое значение для оценки сдвигов, происходящих в энергетическом обмене на ранних этапах трансформации, детерминированной онковирусом, приобретает анализ структурной перестройки митохондрий, осуществляемый параллельно с определением функциональных изменений, полученных на этой же системе и в те же сроки с помощью биохимических методов. Сведения о подобных исследованиях в литературе отсутствуют. А. И. Агеенко, А. С. Ягубов и Ю….
Преобладание медленно движущихся (катодных) фракций ЛДГ4 и ЛДГ5 при снижении уровня быстро движущихся (анодных) ЛДГ1 и ЛДГ2 обнаружено в тканях, адаптированных к гипоксии и ацидозу, в которых основным источником энергии является гликолиз. В то же время можно считать доказанным, что удельный вес гликолиза в клетках злокачественных новообразований повышается (В. С. Шапот, 1975). Установлено, что скорость…
Показано, что повышение процента ЛДГ4 и ЛДГ5, а также снижение содержания ЛДГ1 и ЛДГ2 наблюдались в тканях у большей части обследованных больных (Р<0,001), не только в гомогенатах злокачественных опухолей, но и в тканях полипов прямой и ободочной кишки. При этом отсутствие увеличения содержания ЛДГ5 в опухоли при раке выявлено только в 8,7% случаев (в 17…
Основной характерной чертой трансформированных клеток является перестройка функционирования генетического аппарата, включающая отклонения в регуляции транскрипции и посттранскрипционных модификаций. Эти изменения обусловливают, в частности, перераспределение изоферментного профиля многих ферментов (А. И. Агеенко, 1974; В. С. Шапот, 1975). Это связано с тем, что изоформы определенных ферментов, являясь функционально однородными, имеют некоторую генетическую разобщенность, т. е. их синтез…