Нуклеиновые кислоты, индуцированные при заражении клеток онковирусами
В экспериментах по молекулярной гибридизации и определению плавучей плотности вновь синтезированной ДНК показано, что в инфицированных паповавирусами клетках происходит именно индукция синтеза ядерной ДНК.
В непермиссивных системах вновь синтезированная ДНК представляла собой обычно нормальную, стабильную клеточную ДНК, которая в этих случаях не дегенерировала в процессе инфекции (Dulbecco, 1976). Следовательно, онковирусы активируют большую часть непермиссивных клеток, которые в соответствующих физиологических условиях должны были бы синтезировать ДНК.
Иными словами, механизм этой индукции, вероятно, близок или аналогичен механизму запускания синтеза ДНК в незараженных клетках.
В продуктивной системе на первом этапе, т. е. до начала синтеза вирусной ДНК, синтезируемая клеточная ДНК также представляла собой нормальную ядерную ДНК. На втором этапе, когда впервые выявляется синтез 20S ДНК вируса PY и SV40, некоторые количества вновь синтезируемой клеточной ДНК характеризовались гетерогенной седиментацией в щелочном градиенте плотности сахарозы: промежуточной (38S ДНК) между константой седиментации нормальной клеточной (105S ДНК) и ДНК (20S) паповавирусов.
Эта ДНК составляла 10 — 15% от общего количества ДНК, синтезированной в инфицированных клетках; ее молекулярная масса на 2,1 X 107 меньше, чем масса ДНК, синтезированной в тех же условиях в незараженных клетках или в клетках при абортивной инфекции. Установлено, что гетерогенно седиментирующая клеточная ДНК не является результатом деградации ДНК, предсуществовавшей в клетках к началу инфекции, либо нормальной клеточной ДНК, синтезированной в любое время инфекции и затем под влиянием каких-либо факторов частично разрушенной.
Оказалось, что на поздних стадиях продуктивной инфекции паповавирусами в клетках синтезируется особая клеточная ДНК, характеризующаяся наличием одиночных разрывов (вероятно, произведенных вирусной эндонуклеазой) во вновь синтезированных цепях и превращающаяся позднее в нормальную клеточную ДНК.
Образование таких укороченных цепей клеточной ДНК может быть и следствием того, что в период синтеза вирусной ДНК клеточная ДНК-лигаза не успевает соединять все вновь синтезированные фрагменты ДНК. Другим видом ДНК, синтез которой также стимулируется при инфекции паповавирусов, цитомегаловирусов человека и RSV, является митохондриальная ДНК. Обнаружено, что EcoRI в этой ДНК из зараженных и незараженных клеток делает один двухцепочечный разрыв (Nass, 1977).
При электронно-микроскопическом анализе гетеродуплексной митохондриальной ДНК из инфицированных и незараженных клеток гетерогенность в нуклеотидном составе этих ДНК не выявлена. Наряду с возрастанием синтеза ядерных и митохондриальных ДНК заражение вирусами PY, SV40 и аденовирусами человека 2-го, 7-го и 12-го типов в пермиссивных и непермиссивных клетках сопровождалось усилением синтеза клеточных мРНК.
Однако стимуляция синтеза клеточной мРНК не связана с инициацией вирусиндуцированной репликации клеточной ДНК (May et al., 1976). Установлено, что начало стимуляции синтеза клеточной мРНК совпадает с появлением Т-антигенов, вирусиндуцированной тимидинкиназы, уридинкиназы и вирусной ДНК. Отмечено, что стимуляция касается только синтеза мРНК и не затрагивает синтеза других видов клеточных РНК.
Синтез мРНК гистона продолжается и после снижения синтеза клеточной ДНК. Оказалось, что скорость синтеза мРНК гистона превышает скорость тотального синтеза ДНК (вирусная+клеточная) в зараженных клетках, а не только скорость синтеза ДНК клетки-хозяина (Liberti et al., 1976). Включение 3Н-лизина в гистон f1 не уменьшается и на поздних сроках инфекции, что может свидетельствовать о координированной транскрипции генов гистона.
Предполагают, что стимуляция синтеза клеточных мРНК обусловливается функциями ранних вирусных генов.
Остается только неясным, ускоряют ли эти гены продукцию уже синтезирующихся мРНК или же инициируют транскрипцию мРНК с цистронов, которые до вирусной инфекции были репрессированы.
Очевидно, индуцированная вирусом клеточная ДНК наряду с использованием ее в пролиферативных процессах может утилизироваться и в других целях как вирусом, так и клеткой. Показано, что в состав псевдовирионов включается вирусиндуцированная клеточная ДНК, которая на поздних стадиях инфекции фрагментируется до размеров, соответствующих вирусной ДНК.
Возможно, что вновь синтезированная ДНК используется для репарации поврежденных вирусом хромосом.
Выживание части клеток при высокой множественности инфекции, по-видимому, связано со стимуляцией синтеза клеточной ДНК, которая, вероятно, создает возможность восстановления повреждений хромосом.
«Механизмы вирусного онкогенеза»,
А.И.Агеенко
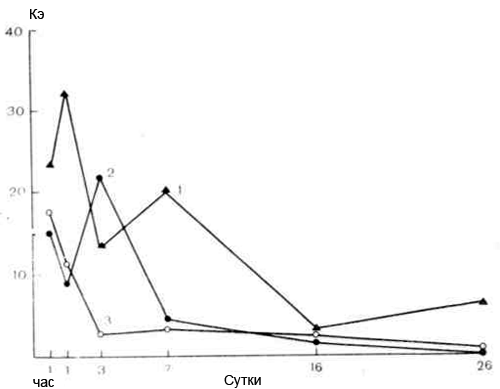
Анализируя результаты, полученные при количественном исследовании тонкого строения клеток ФЭХ при действии вируса А12, можно отметить, что показатель Кэ, характеризующий степень развития эргастоплазмы, значительно снижается, особенно начиная с 7-го дня после инфицирования, и к сроку выраженной трансформации достигает предельно низких величин. Динамика изменения показателя Кэ в процессе трансформации культуры ФЭХ вирусами А12 и RSV 1…
Особое значение для оценки сдвигов, происходящих в энергетическом обмене на ранних этапах трансформации, детерминированной онковирусом, приобретает анализ структурной перестройки митохондрий, осуществляемый параллельно с определением функциональных изменений, полученных на этой же системе и в те же сроки с помощью биохимических методов. Сведения о подобных исследованиях в литературе отсутствуют. А. И. Агеенко, А. С. Ягубов и Ю….
К принципиально важным вопросам онкоморфологии относятся выявление и определение сходства и отличий нормальных и опухолевых клеток. Это стало возможным только в последние годы благодаря количественной электронной микроскопии, основанной на морфометрическом анализе электронно-микроскопических картин с использованием методов планиметрии и принципов стереологии (Г. Г. Автандилов, 1973). Установлено, что эргастоплазма и полирибосомный аппарат являются основными цитоплазматическими органеллами белкового…
Преобладание медленно движущихся (катодных) фракций ЛДГ4 и ЛДГ5 при снижении уровня быстро движущихся (анодных) ЛДГ1 и ЛДГ2 обнаружено в тканях, адаптированных к гипоксии и ацидозу, в которых основным источником энергии является гликолиз. В то же время можно считать доказанным, что удельный вес гликолиза в клетках злокачественных новообразований повышается (В. С. Шапот, 1975). Установлено, что скорость…
Показано, что повышение процента ЛДГ4 и ЛДГ5, а также снижение содержания ЛДГ1 и ЛДГ2 наблюдались в тканях у большей части обследованных больных (Р<0,001), не только в гомогенатах злокачественных опухолей, но и в тканях полипов прямой и ободочной кишки. При этом отсутствие увеличения содержания ЛДГ5 в опухоли при раке выявлено только в 8,7% случаев (в 17…