Ствол мозга
Чередование вдоха и выдоха обусловлено активностью нейронов, расположенных в варолиевом мосту и продолговатом мозге. Считается, что именно здесь находятся дыхательные центры. Они представляют собой не отдельные ядра, а довольно диффузные скопления нескольких групп нейронов.
В стволе мозга различают три основные группы дыхательных нейронов
I. В ретикулярной формации продолговатого мозга располагается медуллярный дыхательный центр. Он состоит из двух отдельных зон. Первая из них включает в себя группу нейронов, локализованных в дорсальных отделах продолговатого мозга (дорсальная дыхательная группа) и активизирующихся главным образом при вдохе. Вторая зона располагается в вентральных отделах продолговатого мозга (вентральная дыхательная группа) и связана преимущественно с выдохом.
Довольно распространено (хотя и не общепринято) мнение о том, что нейроны инспираторной зоны способны к самопроизвольному периодическому возбуждению, и именно они отвечают за периодичность дыхания. При устранении всех возможных афферентных стимулов эти инспираторные нейроны продолжают в определенном ритме генерировать залпы потенциалов действия, которые передаются к диафрагме и другим инспираторным мышцам.
Очередное возбуждение нейронов инспираторной зоны начинается после латентного периода (т. е. периода отсутствия активности) длительностью в несколько секунд. Затем появляются потенциалы действия, и частота их в следующие секунды экспоненциально увеличивается. Соответственно этому нарастает и активность инспираторных мышц. Затем генерация потенциалов действия в инспираторной зоне прекращается, и тонус этих мышц снижается до исходного уровня.
Нарастание импульсации от инспираторных нейронов может быть прервано тормозящими импульсами от пневмотаксического центра. При этом вдох будет укорочен, и в результате возрастает частота дыхания. Кроме того, активность инспираторных нейронов модулируется сигналами, поступающими по блуждающему и языкоглоточному нервам. Эти нервы оканчиваются в одиночном тракте продолговатого мозга, расположенном рядом с инспираторной зоной.
При спокойном дыхании активность экспираторной зоны не проявляется: мы уже знаем, что в этих условиях вентиляция обеспечивается активным сокращением инспираторных мышц (преимущественно диафрагмы), а затем — пассивным возвратом грудной клетки к исходному состоянию. Однако при форсированном дыхании (например, при физической нагрузке) выдох становится активным в результате активизации экспираторных нейронов. Общепринятого мнения о том, каким образом в медуллярных центрах обеспечивается собственный дыхательный ритм, пока не сложилось.
II. В нижних отделах варолиева моста расположен апнейстический центр. Он называется так потому, что перерезание ствола мозга непосредственно выше этого центра вызывает у подопытного животного длительные судорожные вдохи (апнейзисы), прерываемые кратковременными выдохами. По-видимому, импульсация апнейстического центра возбуждает инспираторную зону продолговатого мозга, удлиняя тем самым время генерирования ее потенциалов действия. Неизвестно, играет ли этот центр какую-либо роль в нормальном дыхании у человека; подмечено лишь, что в некоторых случаях при тяжелых поражениях головного мозга у больных может возникать апнейстическое дыхание.
III. В верхних отделах варолиева моста расположен пневмотаксический центр. Как уже говорилось, его импульсы способны подавлять вдох, регулируя глубину и, следовательно, частоту дыхания. Это было показано в опытах на животных с прямой электростимуляцией пневмотаксического центра. Некоторые исследователи полагают, что он связан лишь с «тонкой настройкой» дыхательного ритма, поскольку нормальный ритм может сохраняться и в отсутствие данного центра.
«Физиология дыхания», Дж. Уэст
Дыхание происходит в значительной степени осознано, и в определенных пределах кора головного мозга может подчинять себе стволовые центры. Путем гипервентиляции нетрудно добиться снижения РCO2 в артериальной крови вдвое, хотя при этом возникает алкалоз, иногда сопровождающийся судорожными сокращениями мышц кистей и стоп. При таком снижении РCO2 рН артериальной крови повышается примерно на 0,2. Произвольную гиповентиляцию легких…
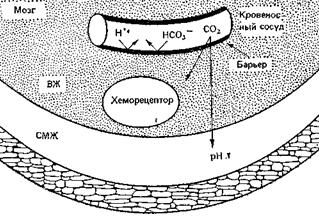
Хеморецепторами называются рецепторы, реагирующие на изменение химического состава омывающей их крови или иной жидкости. Важнейшие из них, участвующие в постоянном контроле вентиляции, расположены у вентральной поверхности продолговатого мозга около выходов IX и X черепно-мозговых нервов. Местная обработка Н+ или растворенным СO2 этой области через несколько секунд вызывает у животных усиление дыхания. Когда-то считалось, что СO2…

Периферические хеморецепторы находятся в каротидных тельцах, расположенных в области бифуркации общих сонных артерий, и в аортальных тельцах, залегающих на верхней и нижней поверхностях дуги аорты. У человека наибольшую роль играют каротидные тельца. В них содержатся две или несколько разновидностей гломерулярных клеток, интенсивно флюоресцирующих при специальной обработке благодаря содержанию допамина. Когда-то считалось, что именно эти клетки…
Существуют три типа рецепторов легких Легочные рецепторы растяжения Полагают, что эти рецепторы залегают в гладких мышцах воздухоносных путей. Они реагируют на растяжение легких. Если легкие длительно удерживаются в раздутом состоянии, то активность рецепторов растяжения изменяется мало, что говорит об их слабой адаптируемости. Импульсация от этих рецепторов идет по крупным миелиновым волокнам блуждающих нервов. Основной ответ…
С дыханием связано еще несколько типов рецепторов Рецепторы носовой полости и верхних дыхательных путей В носовой полости, носоглотке, гортани, трахее находятся реагирующие на механические и химические раздражители рецепторы, которые можно отнести к описанному вышеирритантному типу. Раздражение их рефлекторно вызывает чихание, кашель и сужение бронхов. Механическое раздражение гортани (например, при введении интубационной трубки при плохо проведенной местной…