Нейрон, синапс и глия как материальные носители энграммы памяти
Нейрон и синапс. Рассмотрим нейрон как элементарную единицу нервной системы, способную к обучению и хранению информации.
Общая модель любого нейрона, по представлениям J. Eccles и П. К. Анохина, имеет вход и выход и обладает проводимостью и способностью к интеграции.
Основные этапы проведения и интеграции нервного импульса (а)
и модель межнейронального взаимодействия (б)
 и модель межнейронального взаимодействия (б)")
Компоненты:
1 — входа (сенсорные фильтры);
2 — интеграции;
3 — проведения;
4 — выхода.
Вход представлен постсинаптическими мембранами сомы нейрона и его дендритов. Конечным результатом прохождения нервного импульса является изменение конформации макромолекул белков мембраны с последующим изменением проницаемости, что приводит к возникновению градуированных возбуждающих или тормозных постсинаптических потенциалов. Последние, как известно, отражают соответственно локальную де или гиперполяризацию мембраны. Универсальным механизмом интегративной деятельности нейрона является конвергенция в нем возбуждений, различных по происхождению и модальности [Анохин П. К, 1975].
В качестве анатомического субстрата, обеспечивающего интеграцию, ранее рассматривался аксонный холмик (нейритный конус), в котором в результате пространственно-временной суммации мембранных возбуждающих и тормозящих постсинаптических потенциалов (ВПСП и ТПСП) изменялся уровень возбудимости с последующим возникновением потенциала действия или его торможением.
По современным представлениям, в его состав входят и сложные ультраструктурные образования нейрона (система микротрубочек, нейрофиламентов, субсинаптических цистерн, шипиковый аппарат и система сократительного аппарата), в которых протекают не изученные полностью процессы формирования потенциала действия. Следует подчеркнуть непрерывность этих структур, начинающихся от пресинаптического окончания одного нейрона и в виде синаптических тяжей, доходящих до постсинаптической мембраны другого нейрона.
Проводимость нейрона обеспечивается аксоном, который проводит потенциал действия со значительной скоростью и без декремента по принципу все или ничего. Аксон осуществляет также активную транспортировку в обоих направлениях белков, ферментов, нейрогормонов и медиаторов, выполняя функцию доставки веществ, необходимых для локального синтеза РНК и белков в его пресинаптических терминалях, а также соединений, необходимых для функционирования синапсов. Проведение потенциала действия в конечную зону пресинаптического окончания (выход) сопровождается высвобождением мобильного пула нейромедиатора, который, проходя через синаптическую щель шириной 20—30 нм, вызывает локальное изменение мембранного потенциала.
Приняв условно, что кодом закрепленного в памяти поведенческого акта является система межнейрональных связей, рассмотрим основные принципы их взаимодействия.
Основные типы возможных взаимодействий нейронов в ЦНС

1 — принцип конвергенции;
2 — дивергенции;
3 — реверберации;
4 — принцип распределения возбуждения по многочисленным нейронным цепям;
5 — принцип выбора оппеде ленного пути проведения через случайные многочисленные связи;
6 — возвратное TOD можение по типу Рэншоу;
7 — пресинаптическое торможение;
8 — латеральное TODMO жение для контрастного усиления рецепторных полей нейронных сетей при их последовательном соединении.
Принцип конвергенции означает, что на отдельном нейроне сходятся афферентные импульсы от сотен других нейронов.
«Нейрохимические и функциональные основы долговременной памяти»,
Ю.С. Бродкин, Ю.В. Зайцев
Как показывают электрофизиологические данные, каждый нейрон может принимать участие в различных реакциях животного, т. е. участвовать в работе нескольких нейронных ансамблей. В то же время он обладает чрезвычайной избирательностью в проведении возбуждения по различным каналам связи. Что же лежит в основе этой избирательности? Как отмечалось выше, нейрон является полицептивным образованием, т. е. он чувствителен к…
Роль глиальных клеток в некоторых протекающих в ЦНС процессах, например обучения и памяти, окончательно не выяснена. По основным физиологическим свойствам своей мембраны они существенно отличаются от нейронов. Так, глиальные клетки пассивно реагируют на электрический ток, их мембрана генерирует не распространяющийся импульс, а градуальный медленный потенциал. Например, при раздражении нейронов коры больших полушарий у кошек регистрируется…
Существуют ли прямые доказательства участия глии в процессах длительного хранения энграммы и в какой степени они определяются нейроноглиальными взаимодействиями? Наиболее убедительными представляются данные В. В. Дергачева (1977) об избирательном нарушении долговременной памяти у крыс, обученных с положительным подкреплением, при введении им в желудочки мозга антиглиального гаммаглобулина. Он же в своей монографии ссылается на данные L….
Наиболее полно роль глии в механизме долговременной памяти представлена в гипотезе В. В. Дергачева, названной им нейроселективной концепцией памяти. Автор постулирует ведущую роль функциональной ДНК в активации преформированных в эмбриогенезе и раннем онтогенезе клонов клеток, которые получили название нейроноглиальных комплексов, специфических для данной информации. В их активации и заключен смысл ее кодирования. Однако данная гипотеза…
Согласно концепции W. R. Adey, нейрон головного мозга обрабатывает информацию двумя параллельными способами: во-первых, широким использованием волновых процессов и, во-вторых, определенной последовательностью нейрональных разрядов. Установлено, что формирование нейронного ансамбля связано со значительным увеличением синхронности биоэлектрической активности нейронов в определенном низкочастотном диапазоне, который лежит в пределах 7—10 Гц, т. е. в полосе тетаритма. Причем такая биоэлектрическая…
Распределение импульсов одного нейрона происходит по различным путям, так как после прохождения импульса между двумя нейронами наступает период отсутствия проводимости между ними, который составляет 150—200 мс. В результате одна и та же нервная клетка, как отмечалось выше, может включаться во множество различных нейронных цепей, участвующих в работе нескольких нейрональных ансамблей. Следовательно, процесс хранения информации в…
Стройную теорию организации лимбико-ретикулярных систем в процессах обучения предложил A. Routtenberg (1972). Процессы организации памяти он трактует с точки зрения реципрокных отношений между системой входа (ретикулярная система) и выхода (лимбическая система). Подтверждением ее явились эксперименты L. Goldstein и J. M. Nelsen (1973) по стимуляции этих систем фенамином, активирующим главным образом ретикулярную формацию, и никотином, возбуждающим…
Функциональной основой процесса сравнения информации является способность гиппокампа регистрировать смещенные во времени возбуждающие импульсы, приходящие из ретикулярной формации, перегородки и ведущие к возникновению тетаритма. Распространение этого ритма в кору головного мозга и повышение степени когерентности между ритмами коры и гиппокампа позволили W. R. Adey (1966, 1977) рассматривать эти процессы как доказательство фиксации следов в этих…
Принцип дивергенции, напротив, предполагает возможность распределения эффекторного потока от нейрона за счет разветвления его дендритов и аксонов и образования множества связей с другими нейронами. Принцип реверберирующих (замкнутых) цепей предполагает существование нейронов с двусторонними обратными связями между собой. Возбуждение в таких системах может поддерживаться относительно длительное время при условии выключения в них тормозных нейронов. Возбуждение распространяется…

Пресинаптическое торможение обеспечивается одновременным возбуждением двух аксонов, один из которых заканчивается синапсом на терминалях другого, последующим уменьшением пороговой величины возбуждения и торможением секреции медиатора. Одним из характерных свойств нейрональных сетей является так называемое латеральное торможение. В этом случае возбуждение, поступающее, например, в нейрон А, передается одновременно и на тормозящие промежуточные нейроны, которые, в свою очередь,…
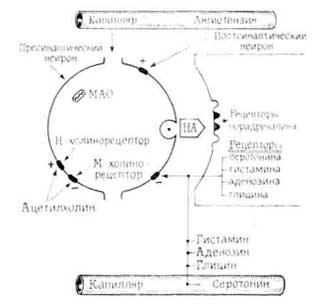
Предполагается, что под влиянием нервного импульса высвобождение медиатора носит квантовый характер. По-видимому, это достигается за счет дискретных изменений в проницаемости пресинаптической мембраны. Гипотеза выделения медиаторов через мембранные каналы более убедительна по сравнению с везикулярной [Marchbanks R. Т., 1978]. Вероятно, везикулы в основном служат резервуаром для хранения запасов медиаторов и их транспорта к пресинаптической мембране. В…
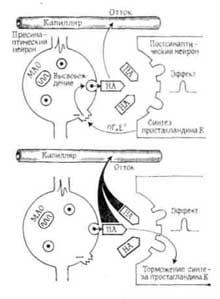
Принято считать, что под воздействием медиатора в области рецептора происходят конформационные изменения его белковых макромолекул. Возникает локальный электрический ток, и мембрана деполяризуется, что вследствие суммарного эффекта сопровождается дальнейшими конформационными превращениями различных ее участков. Поэтому отрицательно заряженная мембрана из гидрофобного состояния, наблюдаемого в покое, переходит в гидрофильное состояние, характерное для возбуждения. Электрический потенциал вдоль таких точек…