Участие пептидных гормонов и малых нейропептидов в процессах фиксации и хранения энграмм
Здесь мы попытаемся изложить современные представления о влиянии пептидных гормонов и некоторых основных малых нейропептидов на процессы памяти человека и животных и обосновать механизмы их действия.
Все пептиды, обладающие специфическим влиянием на собственные механизмы памяти, могут быть классифицированы следующим образом:
Классификация пептидных гормонов и малых нейропептидов, влияющих на поведение и память человека и животных
- Гипоталамические рилизинггормоны и факторы, тормозящие их высвобождение:
- Тиреотропин-рилизинг-гормон (ТРГ).
- Кортиколиберин (КЛ).
- Рилизингфактор лютеинизирующего гормона (РФЛГ).
- Ингибирующий фактор меланоцитостимулирующего гормона (ИФМСГ).
- Соматостатин.
- Пептиды передней доли гипофиза:
- Адренокортикотропный гормон (АКТГ).
- α и β-меланоцитстимулирующий гормон (а и РМСГ).
- Пролактин.
- Фолликулостимулирующий гормон (ФСГ).
- Лютеинизирующий гормон (ЛГ).
- Пептиды задней доли гипофиза:
- Вазопрессин.
- Окситоцин.
- Другие пептиды головного мозга:
- Субстанция Р.
- Нейротензин.
- Опиатные пептиды (β-липотропин, энкефалины, эндорфины).
- Другие пептиды головного мозга:
- Ангиотензин.
- Ренин.
- Гастрин.
- Холецистокинин.
- Нейротензин.
- Опиатные пептиды (Рлипотропин, энкефалины, эндорфины).
В последнее время сложилось представление, что пептидные гормоны, основная роль которых заключается в специфической регуляции гормональных функций, по аналогии с нейромедиаторами вызывают кратковременные или длительные изменения нейрональной возбудимости [Barker J. L., 1976; Hokfelt T. D., 1977]. В частности, Т. D. Hokfelt рассматривает пептиды как возможные нейромедиаторы в ЦНС. Кроме того, в работах ряда исследователей отмечено, что гипоталамические гормоны оказывают влияние на возбудимость нервных структур и поведение, независимо от их влияния на гипофизарноадреналовую систему.
Интенсивное изучение характера влияния этих гормонов, а также гормонов передней и задней долей гипофиза на поведение, высшую нервную деятельность и память за последние 25 лет привело, по сути дела, к рождению новой дисциплины — нейроэндокринологии поведения. Установлено, что большинство пептидов, а также ферментов их синтеза и разрушения локализуется не только в гипоталамусе — основном месте их образования, но и в большинстве других структур головного мозга.
Необходимо отметить, что в спинном мозге найдены локальные вставочные пептидергические нейроны, причем содержание таких пептидов, как субстанция Р, ангиотензин, соматостатин, ТРГ, окситоцин, вазопрессин и энкефалин, в дорсальных рогах спинного мозга выше, чем в вентральных. В значительном количестве пептидергические нейроны обнаружены в желудочно-кишечном тракте.
«Нейрохимические и функциональные основы долговременной памяти»,
Ю.С. Бродкин, Ю.В. Зайцев
Учитывая, что пептидные гормоны существенно изменяют уровень возбудимости нейронов и нейронных популяций в различных образованиях головного мозга, представляло значительный интерес установить зависимость между этими эффектами и влиянием пептидных гормонов на обучение, фиксацию и хранение информации в мозге животных и человека. Больше всего экспериментов проведено по изучению влияния на эти процессы АКТГ. Еще в 1969 г….
Влияние АКТГ на долговременную память также зависит от дозы и времени его введения. Так, по данным R. L. Cold и К. В. van Buskirk (1976), введенный в дозе 0,03—0,3 ME, он увеличивал время сохранения пассивного навыка избегания у крыс, а в дозе 3—10 ME вызывал ретроградную амнезию Стимулирующие эффекты АКТГ независимо от дозы проявлялись сильнее…
Сходное с АКТГ влияние на память и обучение животных оказывает α-меланоцитстимулирующий гормон, структура которого близка к отдельным фрагментам АКТГ: он ускоряет обучение [Paterson А. Т., 1975], значительно увеличивает время хранения закрепленных навыков активного и пассивного избегания электрошока у крыс и облегчает их воспроизведение [Веckwitt В. Е. et al, 1977]. Нужно отметить, что АКТГ, его отдельные…
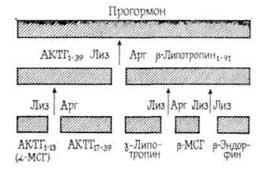
Необходимо отметить, что активным влиянием на память обладают не целые молекулы пептидных гормонов, а в основном их отдельные фрагменты, характеризующиеся специфической последовательностью аминокислот. Следовательно, можно предположить, что эти фрагменты образуются в организме в процессе активного метаболизма пептидных гормонов под влиянием протеолитических ферментов [Чипенс Г. И. и др., 1975]. Интимный механизм пептидных гормонов окончательно не выяснен,…
Совсем недавно Н. Н. Zing и Y. С. Patil (1979) выделили из срединного возвышения и нейрогипофиза крыс два высокомолекулярных белка с относительной молекулярной массой 25 000 и 40 000, обладающих иммунореактивностью кс оматостатину и высвобождающихся при деполяризации нейронов. Авторы предполагают, что один из этих белков является предшественником соматостатина, который синтезируется на рибосомах, высвобождается, а затем…
Многие пептидные гормоны передней доли гипофиза и гипоталамические рилизингфакторы являются субстратами для карбоксиметилтрансферазы, наиболее активной в головном мозге [Diliberto E. J. et al., 1976; Egger D. H. et ai., 1976]. Данный фермент переносит СН3-группу S-аденозилметионина на карбоксильные группы белков, в частности эндогенных акцепторных белков. В результате отрицательный заряд на белке нейтрализуется и становится подвижным (метиловые…

Характер поведения в значительной степени зависит от концентрации пептида на специфических рецепторах. Так, например, рэндорфин при введении его в желудочки мозга крыс в возрастающих концентрациях (0,01 — 0,5 мг) вызывал последовательно возбуждение, замедление угашения условных реакций, обездвиженность подопытных животных и мышечную ригидность [Segal D. S., 1978]. Однако пептиды моделируют также синтез и высвобождение классических нейромедиаторов…
J. D. Вагchas и соавт. (1978) определяют функцию пептидов как нейромодуляторную и предложили соответствующие критерии: Пептиды не действуют транссинаптически. Пептиды действуют только при определенной концентрации в физиологических жидкостях и если имеют доступ к месту возникновения потенциала действия. Эндогенные пептиды должны изменять нейрональную активность постоянным и предсказуемым образом в зависимости от концентрации. Экзогенные пептиды вызывают такой…

Многие из этих пептидов находятся в нервных окончаниях и транспортируются к синапсам с помощью аксоплазматического тока. Субклеточное распределение нейротензина, соматостатина, ангиотензина, субстанции Р, вазопрессина и окситоцина характеризуется их преимущественным связыванием с синаптосомами, выделенными из различных внегипоталамических образований головного мозга [Larner J., 1977; Kobayashi R. M. et al., 1977; Bury R. W. et al., 1977]. Это…
Продукты ферментативного расщепления вазопрессина и окситоцина со специфической последовательностью аминокислот вызывали деполяризацию идентифицированных нейронов гигантской улитки [Takeuchi H. et al., 1977] Тиреотропин-рилизинг-фактор (тиреолиберин) и рилизингфактор лютеинизирующего гормона (люлиберин), а также соматостатин уменьшали спонтанную импульсную активность нейронов коры головного мозга и мозжечка, ствола мозга и гипоталамуса у крыс при ионофоретическом введении. Эти пептидные гормоны уменьшали возбуждение…