Механизм индукции синтеза клеточных нуклеиновых кислот
Прежде всего важно отметить, что онкогенные вирусы способны в обход регуляторных систем клетки активировать всю систему ферментов синтеза ДНК и индуцировать синтез клеточной ДНК, т. е. активировать клеточный механизм, переводящий клетку из G1-фазы в S-фазу.
При этом существенно, что индукция синтеза ДНК вирусом отличается от действия других стимуляторов (например, фитогемагглютинина), которые обычно резко увеличивают синтез всех клеточных РНК, после чего наблюдается репликация ДНК, завершающаяся митозом.
При вирусной же инфекции стимулируется только синтез определенного класса РНК и даже несколько циклов синтеза ДНК может не сопровождаться последующим делением клеток.
Вместе с тем синтезы вирусной и клеточной ДНК не зависят друг от друга, поскольку наличия в ядрах зараженных клеток предшественников и ферментов, обеспечивающих репликацию вирусной ДНК, недостаточно для инициации синтеза клеточной ДНК.
Вероятно, вирусный геном, с одной стороны, обладает собственным инициатором репликации вирусной ДНК, который обусловливает автономность этого процесса и, возможно, осуществляется эндонуклеазой вирусов, а с другой — является специфическим индуктором синтеза клеточной ДНК.
Между двумя этими макромолекулярными синтезами имеется сходство, так как для того и другого необходимы клеточные ферменты, активирующим фактором которых является вирус, а именно ts-A-гкн (это было продемонстрировано на ts-А-мутантах вируса полиомы). Однако функция ts-A-гена необходима только для инициации синтеза ДНК, но не для его поддержания. Нельзя исключить, что репликация вирусной ДНК зависит от физиологического состояния клетки в момент заражения.
Влияние деления клетки на репродукцию вирусов, вероятно, может осуществляться либо за счет конкуренции вирусной и клеточной ДНК за специфические участки, в которых происходит репликация ДНК, либо благодаря тому, что в момент деления клетки создаются условия, приводящие к включению вирусного генома в клеточный.
В том и другом случае будет тормозиться или отсутствовать продуктивная инфекция.
Возможно, активация ферментов, принимающих участие в синтезе ДНК, и последующий синтез клеточной ДНК представляют собой единый процесс, контролируемый одним регуляторным механизмом клетки. Об этом свидетельствует то, что практически вся система ферментов, синтезирующих ДНК, запускается одновременно.
Вирус может, по-видимому, воздействовать «а синтез клеточной ДНК непосредственно через свои продукты, которые дерепрессируют в этом случае определенные опероны и активируют аппарат синтеза ДНК, или опосредованным путем, первоначально повреждая иные системы клетки, приводящие затем к изменению синтеза ДНК.
Мишенью при опосредованном воздействии могут, например, служить клеточные мембраны. Предполагают, что происходящие в процессе трансформации изменения клеточной поверхности как по составу компонентов (полипептидов, гликолипидов, гликопротеинов, появлению вирусспецифических трансплантационных и эмбриональных антигенов), так и по структуре (появлению избыточных рецепторов агглютинина, изменению расположения олигосахаридных цепей и др.) могут служить стимулом для индукции синтеза ДНК и клеточного деления. Особую регулирующую роль при этом отводят кислым белкам, которые синтезируются в цитоплазме в ответ на индуцированные вирусом изменения проницаемости поверхностных мембран клетки.
Учитывая факт одновременной стимуляции ядерной и митохондриальной ДНК, можно предполагать, что вирусспецифический или вирусиндуцировавный фактор, участвующий в этом процессе, способен легко проникать через клеточные мембраны.
В прямых экспериментах с микроинъекциями ранних мРНК SV40 отмечена стимуляция синтеза клеточной ДНК в 80% клетках, синтезировавших Т-антиген (М. Graessman, A. Graessmann, 1976).
Таким образом, эти данные подтверждают гипотезу, согласно которой ранние вирусспецифические мРНК содержат информацию, необходимую для стимуляции репликации клеточной ДНК.
«Механизмы вирусного онкогенеза»,
А.И.Агеенко
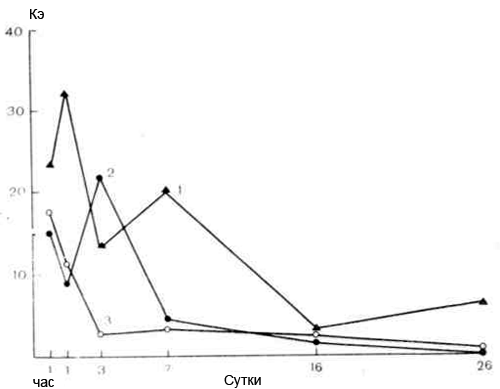
Анализируя результаты, полученные при количественном исследовании тонкого строения клеток ФЭХ при действии вируса А12, можно отметить, что показатель Кэ, характеризующий степень развития эргастоплазмы, значительно снижается, особенно начиная с 7-го дня после инфицирования, и к сроку выраженной трансформации достигает предельно низких величин. Динамика изменения показателя Кэ в процессе трансформации культуры ФЭХ вирусами А12 и RSV 1…
Особое значение для оценки сдвигов, происходящих в энергетическом обмене на ранних этапах трансформации, детерминированной онковирусом, приобретает анализ структурной перестройки митохондрий, осуществляемый параллельно с определением функциональных изменений, полученных на этой же системе и в те же сроки с помощью биохимических методов. Сведения о подобных исследованиях в литературе отсутствуют. А. И. Агеенко, А. С. Ягубов и Ю….
К принципиально важным вопросам онкоморфологии относятся выявление и определение сходства и отличий нормальных и опухолевых клеток. Это стало возможным только в последние годы благодаря количественной электронной микроскопии, основанной на морфометрическом анализе электронно-микроскопических картин с использованием методов планиметрии и принципов стереологии (Г. Г. Автандилов, 1973). Установлено, что эргастоплазма и полирибосомный аппарат являются основными цитоплазматическими органеллами белкового…
Преобладание медленно движущихся (катодных) фракций ЛДГ4 и ЛДГ5 при снижении уровня быстро движущихся (анодных) ЛДГ1 и ЛДГ2 обнаружено в тканях, адаптированных к гипоксии и ацидозу, в которых основным источником энергии является гликолиз. В то же время можно считать доказанным, что удельный вес гликолиза в клетках злокачественных новообразований повышается (В. С. Шапот, 1975). Установлено, что скорость…
Показано, что повышение процента ЛДГ4 и ЛДГ5, а также снижение содержания ЛДГ1 и ЛДГ2 наблюдались в тканях у большей части обследованных больных (Р<0,001), не только в гомогенатах злокачественных опухолей, но и в тканях полипов прямой и ободочной кишки. При этом отсутствие увеличения содержания ЛДГ5 в опухоли при раке выявлено только в 8,7% случаев (в 17…